















Ribozomlar, bakterilerden ökaryotlara kadar tüm canlı hücrelerde protein sentezinin hayati rolünü yerine getiren oldukça karmaşık makromoleküler yapılardır.
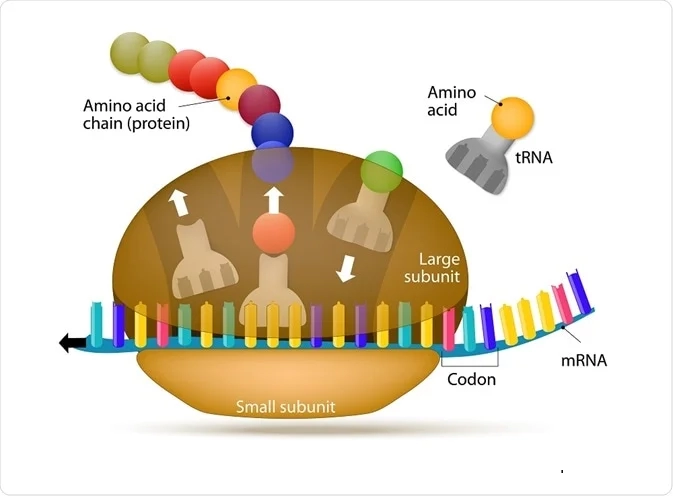
Bir elektron mikroskobu altında görüntülendiğinde, çapları 15 ila 25 nm arasında değişen ribozomlar yassı ve küre şeklinde görünür. Bu yapılar iki ana ribonükleoprotein alt biriminden oluşur. Daha küçük alt birim, kodon adı verilen ardışık üç çerçeve içi baz çifti ile transfer RNA'sı (tRNA) üzerindeki anti-kodon adı verilen tamamlayıcı üçlü arasındaki doğru etkileşimlere aracılık eder.
Daha küçük olanın yaklaşık iki katı büyüklüğünde olan daha büyük alt birim, büyüyen polipeptitteki amino asitler arasında peptit bağının oluşumuna aracılık eden yoğunlaşma reaksiyonunu katalize eden peptidil transferaz merkezini (PTC) içerir. İki alt birim, mRNA'yı çevirmek için bir araya gelir ve çeviri tamamlandığında parçalarına ayrılır.
Ribozomun bölümleri
Ribozomal yapı, tüm türler arasında yüksek oranda korunur, ancak kesin bileşimi organizmaya bağlıdır. Maya, Saccharomyces cerevisiae , ribozomun yapısal dinamiklerinin açıklandığı kanonik modeldir.
Maya, protein ihtiyacını karşılamak için dakikada yaklaşık 2000 üretir. Her ribozom, tek bir işleyen ribozom parçacığı üretmek için gerekli olan dört ribozomal RNA'dan (rRNA'lar) ve 79 ribozomal proteinden (r-proteinler) oluşur; bu nedenle, tüm ribozomların doğru şekilde üretilmesini sağlamak için kayda değer miktarda enerji gereklidir.
Tüm türler arasında, ribozomal alt birimler, Svedberg birimleri ile karakterize edilir, S olarak kısaltılır. Bu birimler, alt birimlerin bir çözücü içinde çökelme hızına dayanır; eşzamanlı olarak, bu, görünen sedimantasyon katsayısı adı verilen bir parametrenin ölçümünden elde edilir.
En basit ribozom bakterilerden gelir ve birlikte 70S ribozomunu oluşturan 30S ve 50S alt birimlerinden oluşur. Buna karşılık, maya ribozomları, 80S yapısını oluşturmak üzere bir araya gelen bir 40S ve 60S alt biriminden oluşur.
Türler arasındaki her bir alt birim, çeviri sürecini destekleyen üç bölge içerir. Bunlar, bir amino asit (aminoasilatlanmış-tRNA) ile hazırlanmış gelen tRNA'yı barındıran A (aminoasil) olarak adlandırılır; tRNA'yı ortaya çıkan veya yeni oluşan peptit zinciriyle (peptidil-tRNA) tutan P (peptidil); ve amino asidinden sıyrılmış (deasile edilmiş-tRNA) ortaya çıkan tRNA'yı harekete geçiren E (çıkış).
Bakteri ribozomlarına göre ökaryotik ribozomlar daha büyük ve daha karmaşıktır. Ek r-proteinleri ve r-protein uzantılarının yanı sıra genişleme segmentleri (ES) içerirler. 79 r-proteinin (diğer ökaryotlarda 80) yanında küçük alt birimde (18S rRNA) ve büyük alt birimde (5S, 5.8S ve 25S rRNA) dağıtılan 5500'den fazla rRNA nükleotidini içerirler.
Tetrahymena thermophila ve Saccharomyces cerevisiae 80S ribozomunun hem küçük hem de büyük alt birimlerinden elde edilen kristal yapıları takiben rRNA'ların ve r-proteinlerin tam ataması ve haritalanması tamamlandı .
Ribozomlar içindeki RNA, yalancı düğümler gibi üçüncül yapısal motifler tarafından düzenlenir ve ribozomun katalitik aktivitesinin yürütülmesinden sorumludur. Proteinler ribozomu dıştan çevreler ve yapısını stabilize ettiğine inanılır.
Serbest ve zara bağlı ribozomlar
Protein sentezinin yeri, proteinin nihai amacına ve hedefine bağlıdır. Protein, onu endoplazmik retikuluma (ER) yönlendiren bir sinyale sahipse, ribozom ER zarına bağlanacaktır. Aynı ribozom, bir sonraki proteinini çevirmeye başladığında parçalara ayrılabilir ve serbest bir ribozom haline gelebilir. Serbest ribozomlar, esas olarak hücre içinde kullanım için proteinleri sentezler. Ökaryotlarda ribozomlar ağırlıklı olarak ER'ye bağlı bulunur.
Bir zara bağlı ribozomlar, hücre dışına ihraç edilmek veya lizozomlarda kullanılmak üzere protein sentezler. ER'de zara bağlı ribozomlar bulunur. Aslında, ribozomların varlığı, ER'ye elektron mikrograflarında imzası olan "kaba" görünümünü verir.
Ribozomun montajı, çekirdekte bulunan bir bölge olan nükleolusta başlar. Burada hem r-proteinleri hem de rRNA'lar için genler aynı anda çevrilir. Translasyonu takiben sitoplazmada r-proteinleri sentezlendikten sonra, sonuç olarak ön-rRNA molekülleri ile birlikte transkripsiyonel bir şekilde birleşir.
Ribozomun muazzam karmaşıklığı nedeniyle, birleştirme işlemi, geçici olarak işlev gören 200'den fazla ribozomal olmayan biyogenez faktörünü kullanır. Pre-rRNA ve r-protein birleşiminden kaynaklanan ilk yapı 90S partikülü olarak adlandırılır; pre-rRNA'nın bölünmesini takiben, 60S öncesi ve 40S öncesi olmak üzere iki parçacığın salınması meydana gelir.
Bu alt birimler daha sonra çekirdekte olgunlaşmaya girdikçe ihracat yetkinliği kazanırlar. Bu olgunlaşma süreci, nükleer gözenek kompleksleri (NPC) yoluyla nükleer zarfı geçtikten sonra sitoplazmada devam eder. Sitoplazmada, daha fazla r-proteinin dahil edilmesi, ribozomal alt birimlerin birleşmeye ve çeviriye başlamasına izin verir.
Olgun ribozomların yapılarına ilişkin önemli bilgiler yapılmış olsa da, ribozomal birleşim süreci, özellikle de kararlı ekspresyon, nükleer ithalat ve r-proteinlerin birleştirilmesinin nasıl gerçekleştiği hakkında daha az şey bilinmektedir.





